Why do tropical songbirds raise only a few chicks at a time?
Tropical songbirds make important life-history trade-offs to maximise the numbers of chicks they produce by producing fewer, high-quality nestlings per breeding effort than temperate zone songbirds
by GrrlScientist for The Guardian | @GrrlScientist

It has been a long-standing ornithological mystery as to why tropical songbirds have smaller clutches of eggs and raise fewer chicks per breeding effort than do temperate songbirds. But today, a study published in the journal Science argues that life history strategies lie at the heart of this conundrum. In this study, evolutionary ecologist Thomas Martin, an Assistant Unit Leader and Senior Scientist at the Montana Cooperative Wildlife Research Unit at the University of Montana, compares nestling growth rates between closely-related species of tropical and temperate songbirds. He documents that nestlings of tropical songbirds grow longer wings, and grow them faster, than do nestlings of temperate songbirds. Further, they achieve longer wings without increasing their time in the nest, which reduces their risk of predation. Further, because tropical songbirds have more resources available to invest into their offspring, they produce fewer chicks per breeding effort and invest more resources into each individual, thereby giving their offspring a higher survival rate after they fledge (leave the nest). In contrast, temperate songbirds have fewer resources available to nurture their chicks and their offspring suffer a higher mortality rate after they leave the nest, so temperate songbirds compensate by producing a greater number of lower-quality offspring.
Life history theory seeks to explain how evolution shapes organisms
Life history theory exists at the intersection between evolution and ecology. It seeks to explain how natural selection shapes the anatomy, physiology and behaviours of living creatures. In the case of reproduction, life history theory examines the trade-offs between the number and size of offspring produced and the intensity of parental effort, juvenile development, age when sexual maturity is reached, age of first reproductive effort, total reproductive lifespan, ageing and death.
Reproduction is the most important and demanding life history event for most individuals. When reproducing, individuals must carefully orchestrate trade-offs so they invest their available resources — time, effort, and energy — into adequately supporting all their life functions in addition to breeding. Since resources are limited, the time, effort, or energy invested into one life function means there are fewer resources available for others.
Practically speaking, life history theory explains why some insects reach sexual maturity in just a few weeks or months, produce thousands of offspring and provide little or no parental care, and drop dead shortly afterwards, whereas humans reach sexual maturity in their teens, produce only a few offspring in their lifetimes, and provide them with extraordinary amounts of parental care throughout the lifetimes of both the parents and the offspring. Thus, life history strategies for all taxa fall somewhere along a fast-slow gradient.

Life history trade-offs are well known for songbirds living at different latitudes. Basically, tropical songbirds occur mostly at the slow end of this fast-slow gradient and northern temperate songbirds are closer to the fast end (doi:10.1111/j.1558–5646.2011.01227.x). Although avian life history strategies have been intensively studied for many decades, no consensus has emerged within the scientific community as to what drives this latitudinal gradient in birds.
Several hypotheses have been put forward to explain the observed latitudinal variations in avian life history strategies. The most widely recognised is the latitudinal gradient hypothesis, which proposes that tropical environments are stable, and contain habitats that are already saturated with individuals and with biodiversity. According to this hypothesis, these are the reasons that the tropics favour slower life history strategies that facilitate improved competitive abilities. But a more exhaustive examination of what wild birds are doing reveals that tropical species actually show more variation along the slow-fast gradient than do temperate bird species, which is inconsistent with this hypothesis (doi:10.1111/j.1558–5646.2011.01227.x). Other hypotheses propose that latitudinal gradients exist for predation risk or for food availability, but they too, do not explain what drives observed latitudinal variations in tropical and temperate birds (doi:10.1111/j.1558–5646.2011.01227.x).
One way to visualise the many life history trade-offs confronted by songbirds is by examining this avian “circle of life” (Figure 1):
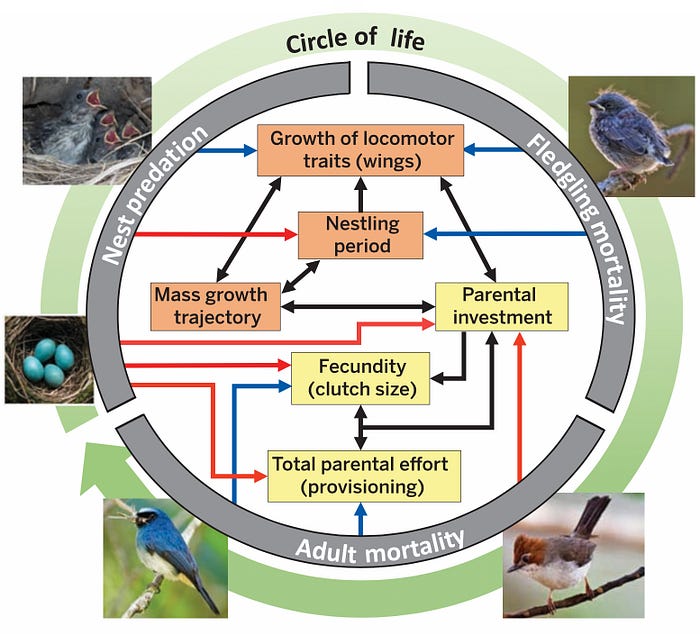
During the different stages in an individual bird’s life, its mortality risk varies (gray ring, Figure 1). This variation in mortality risk exerts selection on growth strategies (orange boxes; Figure 1) and parental strategies (yellow boxes; Figure 1). Selection can either be positive (blue arrows; Figure 1) or negative (red arrows; Figure 1). And of course, one life history trait can influence another (black arrows; Figure 1). For example, nest and fledgling predation exert opposing selection on length of the nestling period.
For many decades, it was assumed that tropical songbird nestlings grow more slowly than temperate songbird nestlings, and thus, it was taken for granted that they also had a longer nestling period. But tropical birds have a high risk of nest/nestling predation, which should select against a longer nestling phase. Further, slower nestling growth does not explain longevity of tropical birds compared with temperate songbirds, so there was no obvious benefit to their slower growth (doi:10.1086/681986).
Clearly, something was just plain wrong here, and that something required a more exhaustive investigation.
For a given level of nest predation, temperate nestlings seem to grow faster
To gain a more complete perspective on daily nest predation rates (Figure 2A), Dr Martin compared predation and growth rates observed in three distinct locations in the field for 20 to 30 songbird species living side-by-side per site. These songbird communities are found in three separate field sites located in north temperate Arizona USA (yellow circles; Figure 2), tropical Malaysia (blue triangles; Figure 2) and tropical Venezuela (black squares; Figure 2):

When Dr Martin compared nestlings with the same body mass, he found that the risk of nest predation is higher for tropical songbirds (Figure 2A). Further, similar trends in growth rates for nestling body mass for all the songbird nestlings examined, tropical songbirds had shorter nestling periods when compared to temperate songbirds with the same body mass growth rates (Figure 2B).
Higher risk of nest predation favours faster nestling growth
To review: overall, tropical songbird nestlings experience similar — or higher — nest predation rates than temperate songbird nestlings yet they appear to have slower growth rates. Yet at the same time, nestling periods for tropical songbirds are 2 to 6 days shorter for the same peak growth rates (Ki) as temperate species (Figure 2B). Which makes no sense.
This made Dr Martin rethink how nestling growth rates are measured and estimated. Traditional peak growth rate estimates (doi:10.1111/j.1558–5646.2011.01227.x) are just that — estimates. Since tropical songbird nestlings have a slower peak growth rate, this was assumed to reflect a slower overall growth. That was a long-standing conceptual error.
When Dr Martin compared the body mass gain over the entire nestling phase for a number of closely-related tropical and temperate songbirds, he found that tropical species (black line; Figure 3) gained weight more slowly and had and a slower peak growth rate than temperate species (yellow line; Figure 3), which showed an early growth spurt. In fact, this paradox embodies the classic tortoise-and-hare scenario. Dr Martin found that nestlings of tropical songbirds showed more sustained growth that actually exceeded that for temperate species after their growth peak (Figure 3). Thus, the tropical nestlings’ growth trajectory gave them a similar average growth rate for the entire nestling period so they ended up with the same body mass as temperate songbird nestlings without extending their vulnerable nestling period (Figure 3):

There were exceptions: Dr Martin reported that, in some cases, tropical songbird nestlings had a slower growth rate along with an extend nestling period (Figure 3H & I), but he notes that those species also have a lower nest predation risk.
“The long-standing perspective has been that tropical birds grow slower overall and this shows that perspective is not correct in many cases”, explained Dr Martin in email. “This is important because it helps resolve the paradox of why tropical birds grow slow when nest predation risk is often high — it turns out many species don’t grow slower overall.”
So where are tropical songbird nestlings investing their energy, if not into increased body mass? Dr Martin suspected this growth pattern may result from tropical nestlings following an alternative life history trajectory. They were investing more energy into wing growth so they could better evade predators — even partially-grown wings assist escape — and also so the young birds could leave the nest with better flight capabilities.
Tropical songbird nestlings grow longer wings faster than temperate species
Dr Martin went back into the field to conduct more studies. He examined the relationships between time spent in the nest, wing growth rate patterns for nestlings, wing size at fledging, rates of parental provisioning, and mortality for both nestlings and parents, and clutch size (Figure 4):


To summarise Figure 4:
- Although relative mass at fledging increases with the length of time spent as a nestling, this does not differ between temperate and tropical songbirds that have the same nestling period (Figure 4A)
- Peak growth rate of wings increases with peak growth rate of body mass, but tropical species have higher peak growth rates for wings when compared to temperate species with the same mass growth rate (Figure 4B)
- Although relative wing size at fledging increases with nestling period among species, tropical species still fledge with relatively longer wings than temperate species (Figure 4C)
- Relative wing size at fledging increases with provisioning rate per offspring while controlling for other factors (Figure 4D)
- Total provisioning rate decreases with nest predation risk (Figure 4E) and increases with annual adult mortality probability (Figure 4F) across species controlled for each other and for other factors
- The only way that tropical songbirds can increase their provisioning rate per nestling is by producing smaller clutches (Figure 4G)
- A Junco hyemalis nestling begging for food from its parent as it takes its first steps to leave the nest (Figure 4H)
Essentially, Dr Martin not only found that tropical songbird nestlings grow relatively faster than their temperate counterparts, but he also discovered that tropical nestlings end up with relatively longer wings by the time they fledge.
“This wing growth is facilitated by high food delivery per offspring in the tropics, which is achieved by having fewer young there. Fewer young is necessary to achieve the higher food delivery per young because parents are constrained in food delivery by offspring and adult mortality”, explained Dr Martin in email.
These novel insights provide a new way to look at an old problem
“For decades, we have evaluated hypotheses, such as nest predation, food limitation, [and] competition, to explain why tropical birds tend to have slow growth rates, live longer, and produce fewer offspring when compared to their temperate counterparts”, said wildlife ecologist Bette Loiselle, director of the Tropical Conservation & Development Program at the University of Florida, who was not part of the study.
“Although attractive, these hypotheses have failed to provide a general explanation for differences across latitudes”, said Professor Loiselle in email.
“The results are clear cut: the main axis of difference between tropical and temperate species is wing development, and that reflects the impact of predation”, said evolutionary ecologist Robert Freckleton, a Professor of Population Biology at the University of Sheffield, who was not part of this study.
Clearly, this breakthrough is based upon upon many years of meticulous field research.

“Tom Martin has invested decades of research to unravel one of the great mysteries in life history evolution. He has amassed extensive and detailed field studies that measure clutch size, parental investment, growth rates, [and] nest predation on large numbers of species at multiple sites”, said Professor Loiselle.
“What is clear from this study is the importance of considering multiple life stages in assessing the evolution of life-history: i.e. it is not just differences in growth rates in the nest that are important, but how these interact with ecological processes, as well as consequences for later development and survival”, agreed Professor Freckleton in email.
“Simply put, Martin has provided an answer to a question that has generated debate for decades. Years from now, we will look back on this paper and cite it as having transformed the way we think about latitudinal differences in clutch size”, said Professor Loiselle. “The importance of extensive field data replicated across species and sites can not be understated — the insights that emerge from this paper to advance our understanding of life history evolution would not be possible without it.”
“These data provide novel insights that lead to a new conceptual framework which uses the risk of mortality at nestling, fledgling, and adult stages to explain variation in life history strategies.”, said Professor Loiselle. “The result is a new way of looking at an old problem.”
When asked what he considers to be the most important feature of this work, Dr Martin replied: “Life histories reflect complex interactions of traits and mortality selection. Consideration of mortality in different life stages, such as the fledgling stage (just after leaving the nest), and traits that might reduce that mortality yields new understanding of parental and growth strategies in both the tropics and temperate zones.”

Source:
Thomas E. Martin (2015). Age-related mortality explains life history strategies of tropical and temperate songbirds, Science, published online on 27 August 2015 ahead of print | doi:10.1126/science.aad1173 [£]
also cited:
Thomas E. Martin, Juan C. Oteyza, Andy J. Boyce, Penn Lloyd and Riccardo Ton (2015). Adult mortality probability and nest predation rates explain parental effort in warming eggs and embryo development time, American Naturalist 186(2): 223–236 | doi:10.1086/681986 (Free)
Thomas E. Martin, Penn Lloyd, Carlos Bosque, Daniel C. Barton, Atilio L. Biancucci, Yi-Ru Cheng, and Riccardo Ton (2011). Growth rate variation among passerine species in tropical and temperate sites: an antagonistic interaction between parental food provisioning and nest predation risk, Evolution, 65(6):1607–22 | doi:10.1111/j.1558–5646.2011.01227.x [£]
.
Love science? Me too! Follow me on Medium for more like this.
.. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..
GrrlScientist is very active on twitter @GrrlScientist and you can follow all her writing by subscribing to her TinyLetter
Originally published on The Guardian on 27 August 2015.
